
هل بمكن القبول بنظرية التطور الصغري ورفض نظرية التطور الشامل؟ هل يمثل هذا حلا وسطا بين رافضي التطور و مؤيديه؟ سنتجاهل الملامح الغير علمية للنماذج البديلة التي تقدم ، ونجادل بأن أي شيء عدا سلف مشترك رئيسي سيؤدي إلى تجزئة تعسفية وغير منطقية لشجرة الحياة، وإلى نظرية للتحدر لا تتفق مع المنطق كما تتعارض مع الأدلة.
عالم النبات السويدي كارل لينيوس، الذي اخترع التسميات البايولوجية للكائنات الحية التي لا تزال تستخدم حتى اليوم، كان خلقياً، حيث يقول: “إننا اليوم نحسب أنواعا بنفس عدد الأشكال المختلفة التي خلقت في البدء”. لقد قامت أعمال لينيوس وأفكاره على فكرة التصميم الإلهي. ولذلك فقد كانت الأنواع ثابتة، وقراباتها الإحصائية تعكس خطة إلهية. ورغم إيمانه بثبات الأنواع، فقد أقدم لينيوس على القبول بحقيقة وجود اختلافات في النوع الواحد. وعام 1756، عند نهاية حياته، أصبح يصدق بأن عدد الأنواع في الجنس الواحد قد يزداد. لم يكن هذا هو التطور كما نعرفه اليوم، لأن لينيوس كان يعتقد بأن الأنواع الجديدة ما هي إلا كائنات مهجنة من الأنواع الموجودة. ولكن ذلك قد أدى إلى أن فكرة ثبات الأنواع أصبحت أقل ثباتاً.
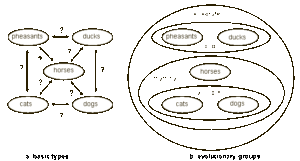
أما عالم الطبيعة الفرنسي جورج لويس بوفون فقد دافع عن ثبات الأنواع في أوائل مسيرته، ولكن في عام 1766، أي قبل حوالي قرن من نشر كتاب أصل الأنواع لداروين، أصبح يقبل بفكرة أن أنواع لينيوس المترابطة فيما بينها بشكل وثيق قد تفرعت من سلف مشترك. وهذا الاعتقاد يشبه إلى حدٍ كبير ما يعرف اليوم بالتطور الصغري. وقد بالغ بوفون في ادعاءاته حتى قال بأن الفصائل التصنيفية هي من خلق الله. تمتلك الفصيلة صفات ثابتة ولكن ليس لها أسلاف. واليوم يجادل الفيلسوف بول نيلسون، وهو من أعضاء حركة التصميم الذكي، لصالح خلقية الأنواع الأساسية (الأصناف الجذرية) النابعة من أسلاف مشتركة. وهو يوضح الأنواع الأساسية بشكل مقتبس من يونكر وشيرر (الشكل 1-أ، 32).
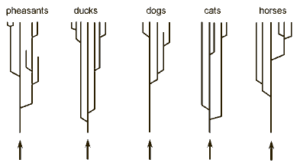
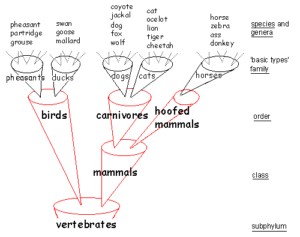
الشكل 1-ب يوضح خمسة مجموعات حيوانية وهي: التدرُج، البط، الكلاب، القطط، والخيول. كل مجموعة انحدرت من سلف مشترك مخلوق بلا سلف مشترك. استبدل نيلسون نظرية الخلق الثابتة (خلق الأنواع الثابتة) بنظرية الخلق الدينامي التي يفضلها. وقد انتقد نيلسون مارك ريدلي (21) لأنه عرض فقط نظرية الخلق الثابتة للأنواع وتجاهل نظرية الخلق الدينامي الحديثة (الشكل 2)
يستخدم نيلسون أربع مصادر لاتهامه ريدلي، منها مصدر الشكل 1-أ ( يونكر وشيرر 1988)، وكلها ظهرت بعد كتاب ريدلي الذي يعود لسنة 1985. وهذا النقد في غير محله، لأنه من البديهي أن ريدلي لن يعلم ماذا سينشر لاحقاً. ليس نيلسون هو الخلقي الوحيد الذي اقترح مثل هذا النموذج. فعند جوناثان صرفتي (26/p.38-39) أيضا نظرية مشابهة تدعى “الكرمة الخلقية الحقة” وذلك لأن شكل هذا النموذج يشبه شكل كرمة العنب. يقترح هذا النموذج أن التنوع حدث في الفترة الزمنية لسفر التكوين. ليس هناك أي أسماء على أشجاره، ولذلك لا يمكن التوصل لأي استنتاج حول المرحلة التصنيفية التي يقصدها. وصرفتي أيضاً يقارن نموذجه بـ “الحشيش” الخلقي.
فنموذج الخلق الأصلي (نظرية الخلق الثابت) يدّعي ثبات الأنواع منذ زمن الخلق، لكنه يقبل بالانقراض. ولكن هذه صورة ساخرة عن فكرة الخلق، وفق رأي صرفتي، لأنها تلمح إلى أن الأنواع التي خلقت في سفر التكوين هي نفسها الأنواع الموجودة حالياً. ويدعي صرفتي أن نموذج الأنواع الثابتة ما هو إلا رجل قش بالنسبة للخلقيين (يقصد هنا أن التطوريين يهاجمون نظرية الخلق الثابتة، غير آبهين بنظريته التي اقترحها). كما يعتقد نيلسون بأن نموذج الأنواع الثابتة لم تتطرق له منشورات وكتب الخلقيين منذ القرن الماضي. وهذا يعني أن الخلقيين لم يتمسكوا بنظرية الخلق الثابتة إلا قبل بضعة عقود. ومن المثير أن نعرف الحقيقة التي سببت تحول الخلقيين إلى نظرية الخلق الدينامي. تشبه نظرية الخلق الدينامية النموذج الذي طرحه الخلقي والتر ريمين (1993، 444 23). فهو مثلا وضع الكلاب، الذئاب، القيوط، ابن آوى والثعالب في مجموعة واحدة تدعى “برامين الكلبيات المفرد”، وهي تطابق نموذج نيلسون للكلبيات. يعرّف ريمين البرامين المفرد بأنه “مجموعة تحوي الكائنات الحية التي تشترك في سلف مشترك واحد” (444). وبالتالي فإن ريمين يقول حتماً: “إن الكائنات المخلوقة مباشرة ليس لها سلف، فقد خلقت مباشرة بالفعل المباشر لمصمم” (510).
تداعيات نظرية الخلق الدينامي:
تستخدم نظرية الخلق الدينامي (DCM) التصور اللاهوتي للخلق كأساس لها: “هنا تعد الأنواع النهائية أعضاء في أصناف أساسية، نابعة من أسلاف مشتركة مخلوقة بذاتها” (نيلسون 2001: 684 16). ولذا فإن DCM هو في أساسه لاهوت. كان بوسع نيلسون أن يهمل كلمة “مخلوق”، وأن يستخدم صياغة لاأدرية مثل “السلف المشترك للفصائل مجهول ولا يمكن للعلم معرفته” لكنه لم يفعل. فلو فعل ذلك فإنه سيحطم أساسات نظرية الخلق. في حديثي التالي سوف أتجاهل الأسس اللاهوتية لـ DCM ليس لأنها غير مهمة أو لان دورها طفيف في المسألة، ولكن لمجرد إبعاد الاتهامات بالإلحاد وتجنب المجادلات والمناقشات التي لا تنتهي حول ما إذا كان التدخل الفوق طبيعي جزءا مشروعا من العلوم الطبيعية. وفي البداية سأقوم بتفحص النتائج أو المقتضيات لـ DCM ثم أقوم بتقييم النموذج نفسه.
1- تداعيات المستوى التصنيفي للأصناف الأساسية:
رغم أن نيلسون لم يذكر المستوى التصنيفي لمجموعاته، فإن سيغفريد شيرر (1998)، مؤلف الرسم الذي استخدمه نيلسون، أوضح أن “الأصناف الأساسية” هي فصائل. ولذلك فإن لدينا فصائل الطواويس، إلأوزيات (البط)، الكلبيات (الكلاب)، السنوريات (القطط)، والخيليات (الخيول). كل واحد من هذه الأصناف الأساسية يضم أجناسا مختلفة (كجنس الكلاب، الثعالب والذئاب) وكل من هذه الأجناس يتكون من نوع واحد أو أكثر. وهكذا فإن الفصائل مخلوقة، خلافا للأجناس أو الأنواع.
2-تداعيات عدد الأنواع الناتجة من الأصناف الأساسية:
تظهر لنا التشجرات mini-trees في الشكل 1-أ فقط 5 – 8 أنواع لكل شجرة. وهذا يعطي انطباعا خاطئا للقارئ؛ فكل جنس يحوي أنواعا أكثر مما صوره لنا هذا الشكل. فمثلاً فصيلة الطواويس تضم أكثر من 12 جنس و 155 نوع، فصيلة البط تضم 41 جنس و147 نوع، فصيلة الكلاب تضم 12 جنس و 34 نوع، وفصيلة القطط تحوي على 37 نوع. وهذه الأرقام لا شيء مقارنة مع فصائل الحشرات: ففصيلة الخنافس (الجُعَل) وحدها تضم قرابة 65,000 نوع (تادج 2000 28). و أحد التداعيات المهمة لذلك هي أن فكرة أن الأصناف الأساسية يجب أن تحتوي كل المعلومات الضرورية لتكوين كل الكائنات الأخرى (سلالاتها المتحدرة منها) غير قابلة للتصديق أبداً. فهذا سيعني، مثلا، أن معلومات 65,000 نوع من الجُعل كانت موجودة في الأصناف الأساسية!! والبديل لذلك هو أن المعلومات الجينية الجديدة تتكون عن طريق الطفرات والانتقاء الطبيعي.
3-تداعيات عدد الأصناف الأساسية:
كم صنفا أساسيا هناك؟ لا يخبرنا الخلقيون. وحتى نعرف ذلك، فإن DCM ليس إلا فتات نظرية. فإذا كانت الأصناف الأساسية قادرة على تضمين ملايين الأنواع على الأرض، فلا بد من وجود الآلاف من الأصناف الأساسية. وحيث أن كل صنف أساسي مخلوق، فهناك الآلاف من أحداث الخلق. وهذه ليست نظرية اقتصادية. أما نظرية داروين للتحدر المشترك فتملك نقطة منشأ واحدة: أصل الحياة.
4-تداعيات عدد “التدخلات”:
في DCM هناك سهم واحد فقط لكل صنف أساس، ولهذا فهناك تدخل خارق واحد لكل صنف أساس. وتداعيات ذلك هو أن سائر تفرعات الشجرة خالية من التدخلات الخارقة. ولو ادعي وجود تدخلات خارقة أخرى، فلن يعود صحيحا أن يدّعى بأن الخلق قد حدث عند مستوى الفصيلة. وبالفعل، فإن تدخلات إضافية كهذه قد تنتهي بالخلق الخاص للأنواع. والخلق الخاص بالأنواع هي جزء من نموذج الخلق الثابت، الذي يرفضه نيلسون قطعا. لا ينكر منظرو التصميم الذكي أن هناك عمليات طبيعية غير موجهة، بل يدعون أن العمليات الطبيعية ليست كلها غير موجهة. ولذا فإن داعمي DCM لا يسعهم الاعتراض على الاستنتاج القائل بأن التشجرات، بغض النظر عن أصلها، هي سلاسل متصلة من العمليات الطبيعية.
5-تداعيات التطفر والانتقاء الطبيعي:
لو حدث الخلق عند مستوى الفصيلة، فلا بد أن الأجناس والأنواع قد نتجت عن عمليات طبيعية. كيف؟ بما أن DCM يدعي أن الأصناف الأساسية تتفاوت (ضمن حدود) بفضل عمليات التطور الصغري، فلا بد أن تكون آلية الاختلاف هي الطفرات والانتقاء الطبيعي الدارويني القياسي. وهذه الآليات أنتجت كل الأنواع “النهائية”. بعبارة أخرى: فالأجناس والأنواع قد نتجت عبر عمليات طبيعية كما تصفها الكتب الدراسية. ولكن من غير المنطقي إذن أن نتحدث عن الطفرات الموجهة وما شاكل. وهناك تداعيات أبعد هي أن من غير المنطقي بعد الآن الاعتراض على فعالية الانتقاء الطبيعي، أو أن الادعاء بأن الانتقاء الطبيعي مجرد حشو أو لا يفسر شيئا (نيلسون 2002: 128 17). وتداعيات أخرى هي أن الرفض المطلق لآليات تكوين الأنواع الجديدة لا يتوافق مع DCM .
6-تداعيات مفهوم التنوع:
في DCM، تعد الأنواع تنويعات على الأصناف الأساسية. ولكن استخدام مصطلح (التنوع) غير مناسب. فالتنوع ظاهرة تحدث ضمن النوع أو المجموعة (شتريكبرغر 2000: 657 27). ومن المضلل استخدام مصطلح التنوع لوصف تشكيل أنواع جديدة بما فيها العوائق الإنجابية. فالعزل الإنجابي هو ما يحافظ على استقلالية الأنواع. ولكن الخلقيين يفضلون مصطلح التنوع للتعبير عن أنه لم يحدث شيء مهم منذ خلق الأصناف الأساسية. كلا اللفظين: الأصناف الأساسية أو الأرضية، لا يوجدان في الكتب الدراسية. وهما يعملان جيدا مع التنوع: الأصناف الأساسية مع التنوع. لكن كلا منهما لفظ غير ملائم.
7-تداعيات التطور الصغري والشامل:
في كتاب دراسي واحد على الأقل (شتريكبرغر 2000: 648 27) يعرّف التطور الصغري بأنه التغيرات التي تحصل ضمن النوع. والتطور الشامل هو فوق مستوى النوع (الجنس، الفصيلة، الرتبة، الشعبة). ووفقاً لـ DCM، فإن تغيرات جديرة بالاهتمام، ولو كانت محدودة، قد تحدث بعد خلق الأصناف الأساسية (نيلسون 2001ب: 684 16). ولكن ما هو مقدار التغيرات؟ وبما أن الجنس أعلى من النوع، فإن DCM يفترض ضمناً حدوث عمليات تطورية شاملة.
8-تداعيات معدل التطور:
إن الجمع بين القول بأن عمر الأرض 6000 سنة، وعدد الأنواع الناتجة من الأصناف الأساسية، ينتج عنه معدل سريع لتشكل الأنواع: 65,000 نوع من الجُعَل خلال 6000 سنة، مما يعني أكثر من 10 أنواع كل سنة!
9-تداعيات تطور الإنسان:
هناك نوع من الكائنات الحية غائب من الشكل 1-أ. ماذا يستلزم DCM حول أصل البشر؟ في التصنيف التقليدي، كان البشر فصيلة مستقلة (البشرانيات). وفي التصنيف الحديث القائم على البيانات الجزيئية، فإن البشر، الغوريلا، الشمبانزي وإنسان الغاب يعودون لنفس الفصيلة (البشرانيات) (فوتويما 1998: 729). ولو حدثت عملية الخلق على مستوى الفصيلة، فإن التصنيف التقليدي سيؤدي لفكرة مريحة هي أن البشر خلقوا منفصلين. وفي التصنيف الحديث، فإن السلف المشترك لفصيلة البشريات مخلوق، وبالتالي فإن البشر، الغوريلا، الشمبانزي، وإنسان الغاب قد تطوروا بطرق طبيعية.
وفي كلا التصنيفين أيضا، فإن البشر، الغوريلا، الشمبانزي وإنسان الغاب جميعا في نفس رتبة الرئيسيات، رُتيبة (أسلاف القردة)، وفيلق (قرود). إذا كان بوسع سلف الكلب أن ينتج فصيلة تحوي على 34 نوع في أقل من عشرة ملايين سنة، فلماذا لم ينتج سلف البشر إلا الشمبانزي، البونوبو، إنسان الغاب، الغوريلا والبشر في نفس المدة؟ علماً بأن التنوع في الصبغيات عند مجموعة أسلاف البشر اقل كثيراً من فصيلة الكلاب (الكلبيات). إذا كان الاختلاف الوراثي (المسافة الجينية) بين الذئب والثعلب مثل ما بين البونوبو والبشر، فعلى الخلقيين أن يخلصوا إلى أن لدى البونوبو والبشر سلف مشترك. ورغم ذلك، فإن الخلقيين يفترضون أن البشر “قد خلقوا عبر فعل مباشر لمصمم.”
10-تداعيات الترتيب النسبي في السجل الأحفوري:
في الشكل 1-أ، كما في الشكل 2، فالمحور العمودي هو محور الزمن. في مخطط يونكر وشيرر، تبدأ الفروع في أزمنة مختلفة، ومن الملفت أن كل الأصناف الأساسية تبدأ في نفس الزمن. فأين الدليل إذن؟ فهو يناقض التسلسل الزمني للسجل الأحفوري، كما أن السجل الأحفوري يظهر لنا أن البكتيريا، أوائل حقيقيات النوى، اللافقاريات، الفقاريات، نباتات اليابسة، الأسماك، الثدييات، والإنسان العاقل لم تظهر في نفس الوقت.
11-تداعيات الزمن المطلق للسجل الأحفوري:
ظهرت فصيلة القطط قبل حوالي 20 مليون عام (شتريكبرغر، 243 27). وتاريخ فصيلة الأحصنة، بما فيها أحافير الخيل، يبدأ من حقبة الإيوسين قبل حوالي 55 مليون عام. وقد تحدرت الكلاب الأليفة من الذئب الرمادي قبل 15,000 ألف عام. وهذه الأزمنة غير متوافقة مع نظرة خلقيي الأرض الفتية الذين يعتقدون أن عمر الأرض هو بين 6,000 و 10,000 عام. وبالإضافة، فإن الأصناف الأساسية المفترض أنها مخلوقة يجب أن تدعمها أدلة من السجل الأحفوري بقدر الأدلة التي تدعم أي سلف آخر في نظرية التطور.
12-تداعيات أصل الأنواع:
هناك نتيجة مهمة للاعتقاد بـ DCM، هي أن كل “المنتجات النهائية”، أي كل الأنواع حالياً، ليست مخلوقة مباشرة. وبما أنه ليس هناك أي “صنف أساس” على قيد الحياة اليوم، فلا يوجد كائن يعيش بيننا اليوم وهو مخلوق مباشرةً. إن زخرفة تدرُج آرجوس Argus pheasant التي وصفها داروين بأنها “أشبه بعمل فني من أن تكون عمل الطبيعة” (داروين 29)، لم تكن من خلق إله، بل نتيجة انتقاء (انظر البندين 4 و 5 من هذه القائمة). فتدرُج آرجوس هو عضو من صنف التدرُج الأساس. وإذا لم يكن السلف المشترك لتلك المجموعة يملك بقعاً عينية على ذيله، فيجب أن تكون الطفرات والانتخاب الطبيعي هي السبب في وجودها. إن طيور الجنة الباهرة في الزخرفة، ذيل الطاووس، ألوان الحمار الوحشي، والدماغ البشري، لا بد أنها قد تطورت عبر الطفرات والانتخاب الطبيعي.
تقييم نموذج الخلق الدينامي (DCM)
الألغاز:
نموذج الخلق الدينامي يستخدم العمليات القياسية للداروينية الجديدة حين يناسبه، ولكنه أيضاً يطرح ألغازاً وتناقضات قاتلة. لنلقي أولا نظرة على الصيغة التقليدية. فالتشجرات تقتضي التحدر المشترك، التطور المتفرع، مستويات التصنيف الهرمية، أصل الأنواع الجديدة، الانتقاء الطبيعي والطفرات. فمثلا، يفسر النموذج التشابهات بين الأصناف الأساسية (كالتشابهات بين الكلاب، الذئاب والثعالب والقيوط) بالتحدر المشترك. وإضافة لذلك، فإن الفروق بين الكلاب، الذئاب والثعالب والقيوط تفسرها الاختلافات بين الكائنات الناتجة من الصنف الأساس السلف. وكلا الحقيقتين تبينهما التشجرات. جيد حتى الآن. ولكن يظهر اختلاف هائل عن التفسير الدارويني القياسي: في الـ DCM، يملك كل من الكلاب والقطط أصلا مستقلا. وهي مجموعات غير مترابطة كلياً. ولا يربط بينها التحدر المشترك. مما يدمر التفسير القياسي (الدارويني) للتشابهات فيما بينها. [نظراً لأن نفس المبدأ تم استعماله فيما سبق، يتساءل الكاتب هنا: لماذا لم يتم استخدام أسلوب التشابهات مرة أخرى في هذه الحالة؟] لا يوفر DCM أي تفسير بديل، مما يقدم لنا لغزاً عميقاً. لا يمكن التشديد كفاية على مدى روعة أن هذا النموذج لا يستطيع الإجابة على أسئلة مباشرة مثل: لماذا يتشارك القطط والكلاب في نفس الخصائص ويوضعون في نفس مجموعة آكلات اللحوم، أو لماذا يصنف التدرُج والبط في نفس مجموعة الطيور. فمن ذا ينكر أن القطط، الكلاب، الدببة وابن عرس يتشاركون في خصائص آكلات اللحوم؟ أما النظرية الداروينية فتفسر تشاركهم لهذه الخصائص بفضل سلف مشترك آكل للحوم، كما تفسر الاختلاف بينهم عبر التشعب من السلف المشترك. ويمكن لسؤال التشابهات أن يكرر مع كل صنف أساس. لا تتوقف التشابهات عند حدود الأصناف الأساسية. بل أن تصنيف لينيوس بأكمله في هذه الحالة سيصبح لغزا عند نموذج DCM.


التضارب
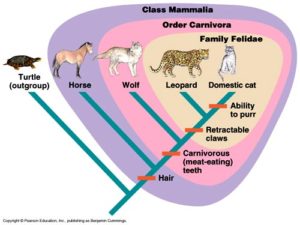
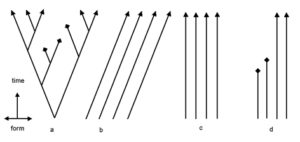
بات من السهل الآن رؤية التضارب المنطقي. فإذا كانت التشابهات والاختلافات سبباً جيداً لتصنيف الكائنات الحية الى صنوف هرمية: أنواع، أجناس وفصائل، ولتفسير هذا النمط بسلف مشترك، فلماذا إذن يتوقف التفسير حين ننتقل إلى الصنوف الأعلى مرتبة كالرتبة، الطائفة، الشعبة والمملكة؟ لماذا يعد السلف المشترك تفسيرا جيدا في مرحلة الفصيلة وسيئا في مراحل تصنيفية أعلى؟ فحتى الخيول والطيور تتشارك في خصائص الفقاريات. وهذا النمط من التشابهات يعرف بنمط المجموعات ضمن مجموعة، أو المجموعات المتضمنة. (الشكل 3-ب؛ الشكل 4). والآن لنعد الى تشجرات نيلسون: فالكلاب مجموعة ضمن اللاحمات، الكلبيات مجموعة ضمن الثدييات، الثدييات مجموعة ضمن القحفيات، والقحفيات مجموعة ضمن الحيوانات. ومنطق المجموعات المتضمنة يجعل من الصعب التصديق بأنها مجموعات منفصلة. فكل مجموعة تصنيفية (ما عدا أعلى رتبة) توجد في مجموعة أعلى منها مستوى. ولا يوجد ما يعرف بمجموعة منفصلة.
مزرعة الحيوان
وفقاً لـ DCM فأن لدى الأصناف الأساسية أصولا مستقلة. وهذا يعني ان الأصناف الأساسية لا يربط بينها التحدر المشترك. كل الأصناف الأساسية متساوية بمعنى أنها مستقلة بالتساوي. وفي محاكاة لجورج أورويل (“كل الحيوانات متساوية، لكن بعضها أكثر تساوياً من بعض”)، فكل الأصناف الأساسية غير متساوية، ولكن بعضها أشد لاتساوياً من بعض. وبالفعل، فهناك درجات من التشابهات. ورغم أن الأصناف الأساسية الثلاثة، طائر التدرُج، البط والقطط، مثلا، لها اصول منفصلة، فإن الخلقيين لا يسعهم انكار أن طائر التدرُج والبط أكثر تشابهاً من طائر التدرُج والقطط. ولكن لماذا تبدو بعض الأصناف الأساسية أكثر تشابهاً من الاخرين؟ ولماذا تبدو بعض الأصناف الأساسية أقل شبهاً بغيرها؟ ولماذا نتوقع وجود تشابهات فوق مستوى الأصناف الأساسية بأي حال؟ فكل نمط من تشابهات الأصناف الأساسية ممكن. ولا يمكن لنموذج الأصناف الأساسية المنفصلة التنبؤ حول نمط المجموعة ضمن المجموعة. فالمجموعات الأعلى مستوى كالطيور، الثدييات، الزواحف، الاسماك والحشرات لا تتوقعها أو تتنبأ بها إطلاقا نظرية الأصول المنفصلة. في الواقع، فإن العلاقات بين الأصناف الأساسية غير معروفة من حيث المبدأ (الشكل 3-أ)، في حين تصبح العلاقات التطورية بين الأنواع أوضح حين تتوفر معلومات جديدة.
إن ما يمكن استنتاجه من النموذج الخلقي الدينامي هو ان وحدة الكائنات الحية مجرد خدعة وتنوعها هو مزحة. يحاول شيرر (1998، 206 25) أن يقدم تعريفا تجريبيا للأصناف الأساسية. لكن الحقيقة هي أنه لم يحاول أبدا التأسيس للتشابهات ودرجات التشابه بين الأصناف الأساسية. كل بايلوجي يصنف التدرُج والبط في مجموعة واحدة، ويفسر التشابهات بينهما بأنها تدل على سلف مشترك أقرب مما بين التدرُج والقطط. وتفسير نظام التصنيف الهرمي اللينيوسي ينهار إذا قبلنا بالأصول المستقلة للأصناف الأساسية المفترضة.

المصدر :- Carl Zimmer (2013) Evolution. Making Sense of Life, page 35
خطة الخلق
في عصر لينيوس، كان وجود نمط المجموعات ضمن مجموعة “يفسر” بأنه خطة للخلق. وقد كان، في الواقع، حقيقة غير مفهومة. فبدون التطور، لا يمكن لأحد أن يأمل في تفسير افضل. وربما لم تكن الحاجة لهذا التفسير محسوسة أصلا. ولكن من غير المقبول أن ننحدر الى أسرار ما قبل الداروينية في حين يوجد لدينا تفسير جيد. فليس من التقدم العلمي فعلا أن تطرح أسرارا ، تناقضات، وتحطّم تفسير كل المجموعات التصنيفية فوق من مستوى الفصيلة. لقد عرض داروين تفسيرا أنيقا لنمط المجموعة ضمن مجموعة، يتمثل بالتحدر المشترك.
فالنموذج الخلقي يحافظ على الغموض والاستعصاء لكل تلك التشابهات التي فسرها التحدر المشترك بكل وضوح وأناقة. فحتى لو قبلنا بالخلق كتفسير علمي فسوف يظل نمط التشابهات دون تفسير. وافتراض الأسلاف المخلوقين لا يقودنا الى توقعات معينة حول أنماط الحياة على الارض مثل نمط المجموعة ضمن مجموعة. وكيف يمكن له؟ فداروين، بعد كل شيء، طرد التكهنات حول “خطة خلق غير معروفة” من العلم (جونسون 1993، 70 10).
إعادة اختراع التحدر المشترك
حاول شيرر (1998 25) أن يعبر عن العلاقات البايلوجية بين الأنواع بصنف نظامي جديد: الأصناف الأساسية. لن يفيد هذا الصنف في تصنيف الأحياء المنتمية لأصناف أساسية مختلفة. فلكي نستطيع تحديد العلاقة بين التدرُج، البط وكل الطيور الأخرى، فنحن نحتاج الى صنف أساسي يدعى الطيور، وهو سلف كل الأصناف الأساسية الأخرى للطيور. ونفس الفكرة تنطبق على كل الثدييات والحيوانات. فنحن نحتاج لأصناف أساسية تدعى “الثدييات” و “الحيوانات”. ولكي نفهم العلاقة بين كل أشكال الحياة على الأرض فنحن نحتاج الى صنف أساس يدعى الحياة. وذلك يكافئ إعادة اختراع السلف المشترك. ويترتب عليه أن الأصناف الأساسية لدى نيلسون وشيرر ليست “أصنافا” ولا “أساسية”. فالطفرات والانتقاء الطبيعي يمكن أن تتجاوز قيود الأصناف الأساسية، متجاهلة كل حدودها.
اختراع داروين
إن انتقاد نيلسون لمخططات ريدلي للتواريخ المختلفة للأنواع على الأرض (ريدلي، 1985) غريب لعدة أسباب. فنموذج الخلق الدينامي يضم العديد من التشجرات. ولكن فكرة استخدام الشجرة لتمثيل العلاقة بين الأنواع مسروقة من داروين وليست بالتأكيد من اختراع الخلقيين.
يقول ماير “كان داروين، بنحو ملفت، أول مؤلف يقترح ان كل الكائنات الحية قد انحدرت من سلف مشترك، عبر عملية مستمرة من التفرع … وقد يكون التزايد المستمر للأنواع تفسيرا لتنوع الحياة العضوية بأكمله” (ماير 1982، 507 14).
وبالإضافة، فالتشجرات في مخطط ريدلي تحتوي على فروعا متباعدة، أما في مخطط نيلسون فهي عمودية. وهذا يعني أن الأنواع ثابتة. ولهذا يعتبر نموذج الخلق الدينامي تشويها للتحدر المشترك. بل لا يمكن لأحد أساساً الادعاء بأن DCM في وسط الطريق تجاه التحدر المشترك. ذلك أن نيلسون وبقية منظري التصميم الذكي جاهلون بقوة وهدف التحدر المشترك. فالتحدر المشترك لا يفسر الفصائل؛ بل الحياة بأكملها. إن وضع حدود لمجال التحدر المشترك هو أشبه بقيادة طائرة على الأرض وتجاهل أنها بنيت لتطير!
أدلة جديدة على التحدر المشترك “للأصناف الأساسية”
حتى الآن كنت اركز في على المنطق والقدرة التفسيرية. ويمكن فهمهما دون معرفة مفصلة بعلم الجينات والكيمياء الحيوية. في الواقع، فإن داروين لم يعلم أي شيء عن الجينات والكيمياء الحيوية، لكنه فهم منطق نظرية التطور. عام 1900 ولد علم الجينات المندلية، وبعدها بخمسين سنة اكتشف جيمس واطسون وفرانسيس كريك تركيب الحمض النووي، وبعد خمسين سنة اخرى نُشِر التسلسل الكامل للجينوم البشري. ويدل الحدث الاخير على تحول علم الجينات (دراسة جينات مفردة) إلى الجينوميات (دراسة الجينوم، أو المجموعة الكاملة لجينات النوع). وحين بدأ العلماء بمقارنة جينومات الأنواع مع بعضها، بدأت أدلة جديدة مذهلة تظهر على التشابهات والتحدر المشترك للأنواع الحية.
فقد كشف التسلسل الجيني للانسان والفأر، مثلا، أن هناك جينات، بل وقطعاً كاملة متطابقة في صبغيات الإنسان والفأر متطابقة. إن قطعة صبغية تتكون من حوالي 90.5 مليون قاعدة دنـا في الصبغي البشري الرابع مشابهة للصبغي الخامس في الفئران. وكل جينات الإنسان تقريباً في الصبغي الـ17 توجد في الصبغي الـ11 عند الفئران. والصبغي الـ20 يعود بأكمله الى القطعة السفلى من الصبغي الثاني عند الفئران. ولرؤية التمثيل الرسومي والتفاعلي لجميع حالات التماثل الجيني بين الفئران والبشر، يمكن زيارة موقع مؤسسة سانجر (2002). وقد اعتمدت الخرائط على بحث (غريغوري وآخرين 2002 9). تعني هذه التشابهات الخاصة أن ما بين المئات والآلاف من الجينات توجد بنفس الترتيب في الإنسان والفئران معاً. وهذا دليل مثير على تحدرهما المشترك.
إن توزيع الجينات على الصبغيات لا يمكن تفسيره بالضرورة البيوكيماوية أو البيولوجية. وباستثناء الصبغي Y وبدرجة أقل الصبغي X، ما من صبغي مكرس لوظيفة حيوية خاصة كالهضم، التنفس، التنقل، التكاثر او الإدراك. فالجينات التي تتحكم بهذه العمليات الحيوية منتشرة على طول 20 زوج من الصبغيات في الفأر و 23 زوج في البشر. وبالتالي، فهذه التطابقات لا يمكن تفسيرها بفضل الضرورة او الصدفة. إن الحوادث التاريخية هي العامل المهيمن الذي أنتج الحجم، الشكل، التركيب، والعدد لهذه الصبغيات. وبغض النظر عن إعادات الترتيب العديدة التي حدثت منذ انفصال الانسان والفأر، فقد ورث الاثنان حوالي 195 قطعة سليمة ومحفوظة من سلفهما المشترك. في نموذج الخلق، يعود الإنسان والفأر إلى صنفين أساسيين منفصلين. ولكن التطابقات في قطع صبغياتهم تظهر لنا أنهما ليسا صنفين أساسين، بل صنفين مشتقين. وقد كشفت نتائج مشابهة في القطط، الفقمات، الابقار، الخيول والجرذان.
وبالفعل، فحين كنت أعمل على المراجعة النهائية لهذا المقال، نشرت مجلة (Science 13 يونيو 2003) إصدارا خاصا مكرسا لشجرة الحياة. وبين أهم المواضيع هو محاولة تمثيل شجرة الحياة بدمج ما لا يقل عن 3000 نوع (بينسي، 2003 19). وبسبب الكم الهائل من المعلومات التي تحويها الشجرة، أشار المحرر بنبرة لا تخلو من الفكاهة: “إن أفضل عرض لشجرة الحياة سيكون بقطر 1.5 متر.” إن أحدث تأكيد مذهل للسلف المشترك يأتينا من مشروع الجينوم البشري. وقد قدم مدير المشروع فرانسيس كولينز وصفا رائعا وواسع الانتشار لهذه الاكتشافات (31).
منذ بدء مشروع الجينوم البشري، تسارعت الاكتشافات المتعلقة بتفرع الانواع إلى حد أمكن معه بناء أشجار نسب جيني مكتملة تقريبا لعشائر مدروسة بعناية، مثل الثدييات. لقد جمعت معلومات تسلسل الجينات من حوالي 182,000 نوع لحقيقيات النواة في قاعدة بيانات بنك الجينات. ولا نحتاج إلا لإضافة معلومات أكثر لبناء شجرة الحياة النهائية بدقة عالية (33).
مايكل بيهي
ليس كل دعاة التصميم الذكي مثل نيلسون. فمايكل بيهي (1996، 176 1) يدعي قبوله بالتحدر المشترك لكل أنواع الحياة:
أعتقد بأن الأدلة تدعم التحدر المشترك بقوة.
وقد كرر هذا التصريح في نشرة اخرى (بيهي 2001، 697 2):
… بما أني أرفض آلية الانتخاب الطبيعي لا التحدر المشترك.
وموقف بيهي محير. فهو لا يصرح بالسبب في قبوله بالتحدر المشترك. وهذه الاقتباسات قد تكون هي كل ما قاله عن التحدر المشترك. وأعتقد أن بيهي لم يدرك عواقب كلامه. فالتحدر المشترك لكل الكائنات الحية على الأرض يعني أن كل الأحياء على الأرض مترابطة فيزيائياً، تاريخياً وجينياً. التحدر المشترك لكل الكائنات الحية يعني أن كل الكائنات ما هي إلا سلسلة واحدة مستمرة من الأسلاف والذرية. التحدر المشترك يعني أن كل كائن يرث جيناته من الجيل الذي قبله مع تعديل بسيط. وهذا يشمل الأنظمة الغير قابلة للاختزال. كل تدخل فوق طبيعي هو خرق للتحدر المشترك، لأنه يعني أن نظاما جديدا غير قابل للاختزال في الكائن الأول يكشف أنه لم يورث عن والديّ، وسيكون من غير المبرر القول “لقد ورثت كل صبغياتي من والديّ عدا النظام غير القابل للاختزال في الصبغي X، الذي يملك أصولا فوق طبيعية.” وبالمثل، فإن بيهي لا يمكنه الادعاء بأن التحدر المشترك صحيح ما عدا في حالة الانظمة الغير قابلة للاختزال، فالتحدر المشترك لا يسمح باستثناءات كهذه. لان هذا قد يعني انتهاك قوانين الجينات. إن علم الجينات أشد مجالات الاحياء دقة ورصانة. وقد أجريت آلاف التجارب الجينية منذ بداية علم الجينات المندلي الكلاسيكي (1900) وبداية علم الجينات الجزيئي (1953). ولم يسبق أبدا ملاحظة ظهور مفاجئ لنظام معقد لا يمكن اختزاله، في حين أن أنواع الطفرات اللازمة للتطور شوهدت بشكل دائم.
فيليب جونسون
فيليب جونسون هو احد قادة حركة التصميم الذكي. ورأيه عن التحدر المشترك واضح جداً في كتابه الشهير”داروين قيد المحاكمة”:
لأن اعتقاد الخلقيين ينص دوما على أن الله قد خلق الأصناف الأساسية، والتي تفرعت من ثم. إن المثال الأشهر للتطور الصغري عند الخلقيين هو نسل آدم وحواء، الذي تفرع من زوج سلفي مشترك لخلق كل الأعراق المتباينة للنوع البشري
(جونسون 1993، 68 10).
وهذه الفقرة مثيرة للاهتمام حقاً. فهي تقترح أن البشر هم “صنف أساس” وقد خلق كذلك. وعبر الانتقال من “صنف أساس” إلى التطور الصغري، يتجنب جونسون التصريح بأن البشر هم “صنف أساس” وبالتالي مخلوقون. ولكن هذا الاقرار يؤشر بشدة إلى أن البشر “صنف أساس”. ولكن لو كانوا كذلك فإن فكرة جونسون عن الأصناف الأساسية أشد تعقيداً من نموذج الخلق الدينامي لدى نيلسون. إن مثال جونسون عن “الصنف الأساس” يقع عند مستوى النوع وبذلك فهو يؤكد ضمناً أن معنى “التطور الصغري” هو التغيير ضمن الأنواع. ولكن إذا كانت هذه هي نظرة جونسون، فهذا يعني أن كل الانواع مخلوقة، والتطور الصغري يسمح له فقط بخلق تعديلات وتكيفات بسيطة ضمن النوع الواحد. ولكن مهما يكن تعريف الصنف الأساس (على مستوى الفصيلة أو النوع)، فالتطور الصغري في هذه الحالة لا ينتج أنواعا جديدة. فكل نموذج خلقي ينتهي بعدد أنواع أكثر من عدد الأنواع التي بدأ بها، سيحتاج الى آلية طبيعية لانتاج أنواع جديدة. وإنتاج انواع جديدة هو تطور واسع كما تنص عليه الكتب الدراسية. ولكن من الصعب على الخلقي أن يعترف بأنه يقبل بالتطور الواسع. وفي فقرة أخرى نرى أن جونسون قد يرغب بالاعتقاد بالخلق الخاص للبشر:
“إننا نشاهد مباشرة أن التفاح يسقط حين يرمى، لكننا لم نلاحظ سلفا مشتركا للقردة الحديثة أو البشر. فما نلاحظه هو أن القرود والانسان متشابهون جسدياً وكيميائياً أكثر من تشابهها مع الأرانب، الأفاعي او الأشجار. إن السلف المشترك القردي هو فرضية ضمن نظرية تسعى لتفسير كيف ظهرت هذه التشابهات الأكبر والأصغر. هذه النظرية معقولة، وخصوصا في الفلسفة المادية، ولكن رغم ذلك قد تكون كاذبة. أما التفسير الصحيح للعلاقات الطبيعية فقد يكون شيئا أشد غموضا بكثير” (جونسون 1993، 67 10).
ومن جديد، فهذه فقرة ملفتة. فهنا يصرح جونسون بأن فرضية أن الانسان والقرد يتشاركان في سلف مشترك معقولة، لكنها رغم ذلك فقد تكون خاطئة . كما يفشل في توضيح إن كان يقبل او يرفض السلف المشترك، ولماذا. إن القول بأن نظرية علمية قد تكون خاطئة ليس بالشيء الجديد، لأن كل النظريات قد تكون خاطئة. وبالتأكيد، فإن الفكرة القائلة بأن التفسير الصحيح للعلاقات قد تكون غامضة، ليست بالسبب الكافي لرفض السلف المشترك. وبنبرة فكاهية يصرح جونسون أن “التحدر مع التعديلات قد يكون فرضية علمية قابلة للاختبار” (66). ولكن اقتراح أسباب غامضة لعلاقات طبيعية ليس “فرضية علمية قابلة للاختبار”. لقد فشلت في رؤية ما دخل الفلسفة المادية بكل هذا. فجونسون لا يقترح تفسيرا غير مادي. وهو بوضوح لا يقبل فرضية التحدر المشترك، لكنه عاجز عن اعطاء سبب جيد لرفضه لها، كما يفشل في تقديم البديل. وهذا هو العلم وفق التفضيل الشخصي.
ويليام دمبسكي
ويليام دمبسكي، عالم الرياضيات لحركة التصميم الذكي، نشر عمله الرئيسي بعد جونسون وبيهي. ولكن رأيه حول التحدر المشترك أقل تحديدا من رأي خلقيين مثل نيلسون وأشد تجاهلا لصحة الأصل المشترك من مايكل بيهي. لقد ذكر البيولوجي التطوري هـ. ألين أور (2002 18) أن حركة التصميم الذكي عادة ما تعترف بأن البشر، الخنازير وزهور البتونيا يربط بينهم سلف مشترك. ولكن منظّرها الرائد ويليام دمبسكي لا يقبل الأصل المشترك دون شروط (دمبسكي 2002: 314، 315 6). فهو يتجاهل نموذج نيلسون الدينامي. ويفشل في تفسير التشابه بين القردة والإنسان مثلا. تنطوي الداروينية على ادعاء تاريخي (التحدر المشترك) وعملية طبيعية (الانتقاء الطبيعي الذي يعمل على تفاوتات عشوائية). ووفقاً للتصميم الذكي، فإن الآلية الداروينية لا يمكنها تحمل ثقل التحدر المشترك. ولذلك يلقي التصميم الذكي بالسلف المشترك تحت الشك، لكنه في نفس الوقت يفسح المجال لأن تكون فكرة السلف المشترك صحيحة، ولو لأسباب أخرى غير الآلية الداروينية (دمبسكي 2002: 315).
إن دمبسكي محق بشأن التمييز بين السلف المشترك لكل الكائنات الحية وآلية التطور، لكنه مخطئ حول العلاقة بينهما. نعم، كلاهما جزء من الداروينية، لكنه ليس محقا في اقتراح أن الانتخاب الطبيعي والتفاوتات العشوائية هي مبررات للسلف المشترك. فقد كان بوسع لداروين أن يتبنى نظريته حول السلف المشترك على أساس التصنيف وحده. كما يُستدل على السلف المشترك من البيانات المستقلة عن آلية التطور. فالأصل المشترك لا يقتضي بالضرورة أي شيء عن الانتقاء الطبيعي، الحيادية، التدرجية، التوازن النقطي أو السجل الأحفوري. إن دمبسكي مصمم على تقويض آلية التطور. وقد أعرب عن أمله في تدمير الأصل المشترك باعتباره نتيجة منطقية لتدمير آلية التطور. وبشكل ملحوظ، وبعد 150 سنة على داروين، لا شيء لدى دمبسكي ليقوله إلا عبارة غامضة:
إن الوقت يسمح كإمكانية نشطة جدا بأن يكون السلف المشترك صحيحا، ولو لأسباب أخرى غير الآلية الداروينية.
هذه الملاحظة الغامضة شبيهة جدا بعبارة جونسون المطلسمة: “عملية ما تتجاوز مطلقا نطاق معارفنا” (جونسون، 155). وبشكل ملحوظ، فقد قال جونسون في وقت سابق في كتابه أن “التكهنات ليست بديلا للدليل العلمي” وأن “داروين، بعد كل شيء، قد نفى التكهنات حول ’خطة غير معروفة للخلق‘ من العلم” (70). لقد فات الأوان على الغوامض بعد 150 سنة من داروين. فلدى البيولوجيين الآن شيء أفضل: يدعى السلف المشترك. لقد نسي جونسون ودمبسكي معاً أن تشارلز داروين لم يعرف أي شيء عن الطفرات، كما لم يعرف أي شيء عن الوراثة المندلية (التي اكتشفت من جديد في 1900) ولم يكن أيضاً على معرفة بالوراثة الجزيئية (التي ولدت عام 1953). والقصد هو أن داروين كان يملك أسبابا كافية لتفسير أنماط التشابه والاختلاف بين الكائنات الحية، حتى دون أي معرفة بعلم الجينات.
إن نجاح تفسير داروين لم يعتمد أصلا على تفاصيل نظريته للتوريث، التي اكتشف لاحقاً أنها خاطئة. فنحن الآن صرنا نعلم أن لغة الجينات لكل الحياة (كيف تترجم الجينات إلى بروتينات) ليست متشابهة فقط وإنما متطابقة عمليا في كل الأحياء، ولدينا تأكيدات وفيرة على الأصل المشترك. ما كان داروين ليتوقع أن الأصل المشترك ستتوفر له كل هذه الأسس الرائعة. الشفرة الوراثية المحددة التي تستخدمها كل الأحياء لترجمة الجينات إلى بروتينات ربما كانت لتختلف بشكل كبير؛ لم تكتشف أي قوانين كيميائية قد تجعل الشفرة الوراثية الحالية مجرد ضرورة. وكل الأصناف الأساسية التي خُلِقت يمكن أن يكون لها شفرة وراثية مختلفة دون أي مشاكل فسيولوجية أو بيئية (كورتهوف 2001).
حيث يمكن للكلاب والقطط أن يملكوا شفرات وراثية مختلفة. كما يمكن للإنسان والقرد يمكن أن يتلكوا شفرات وراثية مختلفة عن بعضهم، لكننا لا نجد هذا. فالسلف المشترك يمكن دحضه بسهولة إذا كانت الكائنات الحية الأشد قرابة متباينة في شفراتها الوراثية (نظريا، يمكن للشفرات الوراثية أن تختلف تدريجياً). ولكن كل الأنواع تملك نفس الشفرة الوراثية. والتفاوتات النادرة والصغيرة في الشفرة الوراثية تفرض على السلف المشترك، وتتبع نمط التوريث مع التعديل. ويمكن بناء شجرات السلالات الجينية على هذه التفاوتات. إن الشفرة الوراثية، التي تترجم الجينات إلى بروتينات، مخزنة في الدي ان اي وعرضة للطفرات. إن الشفرات الوراثية المتفاوتة لها تأثيرات محددة على الكائن الحي. وكل الشفرات الوراثية المتفاوتة الممكنة مدمرة للكائن الحي وخاضعة لضغوط انتخابية قوية. وهذا يفسر سبب ندرة التفاوتات. وبعيداً عن كونها حجة ضد الأصل المشترك، كما يستدل بعض الخلقيين، فهي تعطينا دلائل عن أصل الشفرة الوراثية.
بالإضافة، فإن داروين لم يقم ولم يتمكن من بناء نظرية الأصل المشترك لتفسير وحدة الشفرة الوراثية. لأن وحدة الشفرة الوراثية قد اكتشفت بعد أكثر من قرن على نشر داروين لكتاب أصل الأنواع. وبالتالي فأن الأصل المشترك يفسر بنجاح حقيقة جديدة تماما عن أصل الحياة على الأرض. كل هذه التشابهات لن تتلاشى، مهما تكن ادعاءات دمبسكي حول عدم كفاية آلية التطور. في الواقع، فمن المحال الإتيان ببديل كاملة ومنظم للأصل المشترك. وقد أدت محاولات صياغة بديل طبيعي للأصل المشترك إلى مشاكل وسخافات شديدة. كمثال، فإن فرضية سيناپاثي وشواپه عن الأصل المستقل فيها مشاكل أكبر في تفسيرها لخصائص الحياة. تابع القصة الكاملة حول موقف دمبسكي من الأصل المشترك (2007) في مدونة إبهام الباندا.
هل هناك بديل للتحدر المشترك؟
هل هناك بديل للتحدر المشترك، او هل يمكن وجود تحدر مشترك جزئي؟ إن نموذج الخلق الدينامي (DCM)، مع أنواعه المخلوقة وتشجراته المصغرة، يحطم عالم الأحياْء إلى أجزاء، في حين يوحّد التحدر المشترك كل الكائنات الحية. في الواقع، فإن التحدر المشترك يوحد كل تخصصات علم الاحياء. فنموذج الخلق لا يفسر التشابهات بين الأصناف الأساسية (الكلاب والقطط’)، وذلك نقص خطير، لأن داروين قد وجد بالفعل تفسيراً أنيقاً لأوجه التشابه بين المجموعات التصنيفية. والخلق يقيد الانتقاء الطبيعي والطفرات بطريقة تعسفية. ولذلك فإن نموذج الخلق الدينامي يفشل في أن يكون إطاراً متسقاً ومتكاملاً للتعامل مع البيانات البيولوجية. ولا يمكنه أن يحل محل التحدر المشترك. ولا يمكن أن يفهم إلا بوصفه محاولة لإعادة تقديم أصناف سفر التكوين وليس نتيجة لمحاولة حقيقية نحو تفسير التنوع والوحدة في الحياة.
ورغم الادعاء بأنه نموذج دينامي وحديث، ورغم أنه في الواقع من التطور أكثر مما تصرح به صياغته، فإن DCM لا يعرض أي تقدم ملحوظ بعد بوفون، عالم الحيوان الفرنسي من القرن الثامن عشر. وهو في الأساس رأي قبل دارويني. وحيث أن الطبيعة لم تقم بأي عمل إبداعي فعلي بعد خلق الأصناف الأساسية (بل مجرد تفاوت ضمن حدود)، فهو في جوهره نظرية جامدة. لقد صنف مارك ريدلي النظرية بأنها جامدة، وقد كان محقا على كل حال. إن آراء بيهي عن التعقيد غير القابل للاختزال وكذلك آراء دمبسكي عن المعلومات المعقدة المتخصصة غير كافية لتفسير التشابهات التي نراها بين القطط والكلاب مثلاً. بل إن دمبسكي لم يتبنى بعد رأياً واضحاً حول حقيقة التحدر المشترك. دون شرح لأوجه التشابه، فهناك فجوة حقيقية في نظريات الحياة لديهما. وحتى الآن، لم يخرج لنا أي شخص بتفسير كامل بديل لكل المشاهدات التي فسرتها نظرية التحدر المشترك بكل دقة. ولذلك فمن البديهي أن نقول أنه لا يوجد حالياً بديل للتحدر المشترك. فالتحدر المشترك هو النظرية الوحيدة المنطقية وغير الاعتباطية والمتوافقة مع الأدلة لتفسير أصل الأحياء.
المصادر والمراجع:
- Behe, Michael, 1996, Darwin’s Black Box, New York: The Free Press. Behe has repeated this claim in The Edge of Evolution.
- Behe, Michael, 2001, “Reply to My Critics,” Biology and Philosophy 16:685-709, 697.
- Bowler, Peter, 1989, Evolution. The History of an Idea, Berkeley and Los Angeles, California: University of California Press.
- Dembski, William, 1998, Mere Creation. Science, Faith & Intelligent Design, Downers Grove, Illinois: InterVarsity Press.
- Dembski, William, and Kushiner, James, 2001, Signs of Intelligence, Grand Rapids, MI: Brazos Press.
- Dembski, William, 2002, No Free Lunch, Lanham, Maryland: Rowman & Littlefield Publishers.
- Dobzhansky, Theodosius, 1964, “Biology, Molecular and Organismic,” American Zoologist 4, 449.
- Futuyma, Douglas, 1998, Evolutionary Biology, Third Edition, Sunderland, Massachusetts: Sinauer Associates.
- Gregory, Simon et al, 2002, “A physical map of the mouse genome”. Nature 418, 743-750
- Johnson, Phillip, 1993, Darwin on Trial, Downers Grove, Illinois: InterVarsity Press.
- Korthof, Gert, 2001, Does life look like or unlike evolution? is a review of Walter Remine (1993) The Biotic Message.
- Korthof, Gert, 2002, A Chemist’s View of Life: Ultimate Reductionism & Dissent is a review of Christian Schwabe’s The Genomic Potential Hypothesis
- Korthof, Gert, 2003 Independent origin and the facts of life is a general overview of the theory of independent origin.
- Mayr, Ernst, 1982, The Growth of Biological Thought, Cambridge, Massachusetts: Harvard University Press. See overview of his books.
- Mayr, Ernst, 2001, What Evolution is, New York, NY: Basic Books.
- Nelson, Paul, 2001, “Unfit for survival. The fatal flaws of natural selection,” chapter 10 in Dembski & Kushiner (2001).
- Nelson, Paul, 2002, “The Role of Theology in Current Evolutionary Reasoning,” p. 684, chapter 32 in Pennock (2002).
- Orr, Allen, 2002, in a review of No Free Lunch in the Boston Review, summer 2002 issue.
- Pennisi, Elizabeth, 2003, “Modernizing the Tree of Life”, Science 300 (5626): 1692-1697.
- Pennock, Robert, 2002, Intelligent Design Creationism and its critics, Cambridge, Massachusetts: The MIT Press
- Ridley, Mark, 1985, The Problems of Evolution, Oxford: Oxford University Press.
- Ridley, Mark, 1996, Evolution, second edition, Cambridge, Massachusetts: Blackwell Science.
- Remine, Walter, 1993, The Biotic Message. Evolution versus message theory, Saint Paul, Minnesota: St. Paul Science.
- Sanger Institute web site
- Scherer, Siegfried, 1998, “Basic types of life,” chapter 8 in: Dembski (4) (1998).
- Sarfati, Jonathan, 2000, Refuting Evolution, Green Forest, AR: Master Books.
- Strickberger, Monroe, 2000, Evolution, third edition, Sudbury, MA: Jones and Bartlett Publishers.
- Tudge, Colin, 2000, The Variety of Life, Oxford: Oxford University Press, p. 287.
- Darwin, Charles, 1871, The Descent of Man, and selection in Relation to Sex, Sexual Selection: Birds. PART II. page 92, darwin-online.org.uk, ID=F937, pageseq=109.
“The case of the Argus pheasant is still more striking. (…) It may well be asked, could such artistically-shaded ornaments have been formed by means of sexual selection? (…) and have declared that it was more like a work of art than of nature.” - This essay is an updated and extended version of a chapter in Matt Young and Taner Edis (2004) Why Intelligent Design Fails: A Scientific Critique of the New Creationism. See also that page for Further Reading.
- Francis Collins (2006) The language of God (review)
- This figure is still present in: Junker & Scherer (2001) “Evolution. Ein kritisches Lehrbuch”. Weyel, Gießen. See for a discussion: Martin Neukamm “Die kreationistische Grundtypenbiologie in der Kritik. – Warum es keine empirisch-wissenschaftliche Schöpfungsforschung geben kann“.
- Michael J. Sanderson (2008) ‘Phylogenetic Signal in the Eukaryotic Tree of Life’, Science, 4 Jul 2008.
- Eugene V. Koonin (2009) ‘Darwinian evolution in the light of genomics’, Nucleic Acids Research, 2009, Vol. 37, No. 4 1011-1034. Free Access.
