
تقدم دراسة التسلسلات الجزيئية للبروتينات نموذجاً نستخلص من خلاله أن التشابه الملحوظ بين الجينات الكلية الوجود أو “واسعة الانتشار” Ubiquitous genes لأنواع مختلفة من الكائنات تعني ارتباط النسب بينها.
جوهر الحجة:
الجينات الكلية الوجود (ubiquitous) هي أنواع من الجينات الموجودة لدى كافة أنواع الكائنات الحية و تؤدي وظائف أساسية للكائن و هي غير مرتبطة بوظيفة معينة لدى أنواع معينة ، إذ لا يهم إن كنت بكتيريا أو حيوان أو إنسان أو حتى نبات ليكون لديك هذه الجينات و بكل الأحوال سوف تقوم بالوظيفة البيولوجية الأساسية نفسها للكائن.
و يمكننا أن نجد عدد هائل من الصيغ الموازية لها وظيفياً أي أنه هناك عدد هائل من تسلسلات بروتينية قادرة على تأدية الوظيف البيوكيميائية ذاتها لذلك من الواضح عدم وجود سبب أساسي يجعل الكائنات تمتلك تسلسل متماثل أو حتى متشابه لأنه لا يوجد تسلسل محدد ضروري وظيفياً لأي نوع من الكائنات، كل ما يهم أن يتوافر عدد كافٍ من الصيغ الموازية وظيفياً لأي من هذه البروتينات الكلية الوجود و أي تسلسل بروتيني متماثل بين كائنان مختلفان فإن سببه هو الوراثة بعيداً عن الصدفة المحضة فكلما كان التسلسل أكثر تماثلاً كلما زادت القرابة بين النوعان.
مناقشة الدليل:
تستعمل تسلسلات الحمض الأميني للبروتينات لتثبيت علاقات النسب في شجرة التطور فقد تركزت دراسة الجينات الوظيفية على البروتينات الكلية الوجود أو الـRNA واسعة الانتشار و التي توجد لدى كل الكائنات مما يضمن أن تكون المقارنات مستقلة عن التكوين الظاهري للأنواع. فلنفترض مثلاً مقارنة تسلسل البروتينات لدى كل من الإنسان والشمبانزي اللذان نظراً لتشابه الوظائف والخصائص التشريحية لهما من المتوقع تشابه البروتينات لديهما أيضاً بغض النظر عن قرابة نسبهما و مع ذلك يمكننا أن نستعمل بروتينات أساسية جداً في المقارنة كـ Cytochrome c الذي لا يملك أي تأثير على خصائص الشمبانزي أو الإنسان.
سيتوكروم سي Cytochrome c هو بروتين أساسي كلي الوجود “واسع الانتشار” في كافة الكائنات بما فيها حقيقيات النوى و البكتيريا حيث تتضمن متقدرات الخلايا (الميتوكوندريا) هذا البروتين الذي ينقل الإلكترونات في عملية الأيض الأساسية لعملية الفسفرة المؤكسدة oxidative phosphorylation التي تنتج الطاقة عبر الأكسجين الذي نتنفسه و باعتماد هذا الجين نجد أنه لا يوجد سبب لنفترض أن اثنان من الكائنات الحية يجب أن يمتلكا تسلسلاً بروتينياً متماثلاً أو حتى تسلسلاً متشابهاً إلا إذا كان هاذان الكائنان قريبين بالنسب وجزءٌ من هذا ناجم عن التكرار الوظيفي لتسلسلات و تركيبة البروتين و نعني هنا بكلمة تكرار أن العديد من تسلسلات البروتين تشكل البنية العامة نفسها و تؤدي الدور البيولوجي نفسه و سيتوكروم سي هو مثال جيد على بروتين متكرر وظيفياً لأن العديد من التسلسلات المتباينة تقوم بتشكيل هذا النوع من البروتينات الناقلة للإلكترون ، وهذا التكرار الوظيفي لا يحتاج للدقة في الأداء حيث قد نجد بعض تسلسلات السيتوكروم سي الوظيفية تقوم بنقل أفضل للإلكترون من غيرها.
و أظهرت عقود من أدلة الكيمياء الحيوية أن الكثير من طفرات الحمض الأميني وخاصة بقايا السطح لها أثر بسيط على وظائف البروتينات و بنيتها ، مثال مدهش على ذلك هو سيتوكروم س لأنواع مختلفة من البكتيريا التي لا تملك تشابه فعلي بين تسلسلاتها ومع ذلك كلها تنطوي تحت البنية الثلاثية الأبعاد نفسها وتقوم بتأدية الدور البيولوجي نفسه.
التسلسل الجزيئي لسيتوكروم سي أساسي للحياة والكائنات التي لا تمتلكه لا تستطيع العيش ، فقد أظهرت الدراسات أن بروتين سيتوكروم سي لدى الإنسان و بالرغم من اختلافه عن ذلك الخاص بالخمائر بنسبة 40% إلا أنه يعمل أيضاً في الخمائر (كائنات أحادية الخلية) و ذلك بسبب مسح السيتوكروم سي الأصلي الخاص بالخمائر ، و كذلك أيضاً مع السيتوكروم سي من سمك الطون ، طائر الحمام، الحصان والجرذ من الثدييات و ذبابة الفاكهة من الحشرات فكلها تعمل في الخمائر.
علاوة على ذلك أظهر تحليل وراثي واسع لسيتوكروم سي أن النسبة الأعلى من تسلسل البروتين فيه غير ضرورية لوظيفته في الوسط الحيوي فقط الثلث من أحماضه الأمينية المئة ضرورية لتحديد وظيفته و معظم هذه الأحماض هي متغيرات زائدة أي يمكن استبدالها بعدد كبير من الأحماض المشابهة لها وظيفياً.
الأهم من ذلك قيام البروفسر هيبورت يوكي بدراسة دقيقة استطاع من خلالها التوصل إلى وجود 2.3×10^93 تسلسل بروتيني محتمل لسيتوكروم سي على الأقل بالاعتماد على تحليلات للطفرات الوراثية، ومن هذا المنظور فإن الرقم 93^10 هو أكبر بمليار مرة من عدد الذرات في العالم المرئي و لهذا فإن عدد تسلسلات سيتوكروم سي الوظيفية هو عدد لا حدود له واقعياً و لا يوجد سبب حينها يجعل من كائنان مختلفان أن يملكا تسلسلاً متماثلاً لهذا الجين.
ضمن التحليل الإحصائي العلمي نجد (فرضية العدم Null hypothesis) التي ترى أن هوية حمض أميني غير أساسي في بروتين سيتوكروم سي لدى كل من الإنسان والشمبانزي يجب أن تكون عشوائية و لكننا نعرف من خلال نظرية الأصل المشترك و نموذج شجرة التطور أنهما مرتبطان بشكل كبير و بذلك سنتوقع أن بروتينات سيتوكروم سي عند الإنسان و قرد الشمبانزي يجب أن تكون متماثلة وذلك ببساطة يعود للوراثة و هو ما نجده حقاً.
تأكيد:
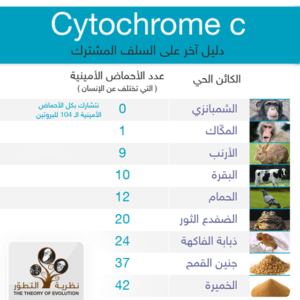
يمتلك الإنسان والشمبانزي التسلسل نفسه لبروتين سيتوكروم سي (نتشارك جميع الأحماض الأمينية الـ104 للبروتين مع الشيمانزي و نختلف مع المكّاك بحمض أميني واحد هو الحمض الـ 66 حيث نملك نحن و الشيمبانزي في هذا الموضع حمض إيزوليوسين I أما المكّاك فحمض ثريونين T) و فرضية العدم السابقة تعتبر خاطئة ففي ظل غياب أصل مشترك فإن فرصة حدوث التماثل هي أقل من 93-^10 أي ( واحد على 10 مرفوعة للأس 93!) و بالتالي فإن الدرجة العالية من التماثل هنا هي تأييد مذهل لنظرية السلف المشترك و علاوة على ذلك نجد أن بروتينات سيتوكروم سي لدى الإنسان والشمبانزي تختلف بـ 10 أحماض أمينية فقط عن باقي الثدييات و فرصة حدوث ذلك في غياب آلية وراثية هو أقل من 29-^10. مثال آخر نجده في خمائر الكانديدا و هي واحدة من أكثر الكائنات حقيقيات النوى بعداً عن الإنسان من ناحية ارتباط النسب إلا أن لها 51 حمض أميني فقط مختلف عن التسلسلات البشرية وتقدير متحفظ عن هذه الاحتمالية يكون أقل من 25-^10.
النقد:
قد نجد اعتراضاً وحيداً ممكناً أمامنا و هو الاختلاف البسيط في الأداء الوظيفي بين السيتوكرومات المتنوعة لكن هذا الاعتراض غير مرجح بسبب العدد الهائل للتسلسلات الموازية لبعضها لدرجة عدم تمييزها من حيث البنية الظاهرية إضافة إلى أن التسلسلات المتماثلة حتى قد تعطي درجات مختلفة من الأداء.
مع ذلك و بقصد الحجة لنفترض أن الكائنات ذات عمليات الأيض النشطة و معدلات الانقباض العضلي العالية قد تتطلب سيتوكروم سي أسرع في نقل الإلكترونات لديها فإن نمطاً ملحوظاً هنا من التشابه في التسلسلات سوف يظهر و لسوف يرتبط هذا مع تشابه في البيئة والمتطلبات الفسيولوجية إلا أن ذلك غير ملاحظ أبداً ، على سبيل المثال سيتوكروم سي لدى الخفاش أقرب لسيتوكروم الإنسان منه لطائر الطنان و سيتوكروم سي الدلفين أقرب للإنسان من القرش.و الشجرة التطورية المبينة على تشابه السيتوكروم سي تشابه تلك القائمة على أدلة أخرى لا علاقة لها بالسيتوكروم سي.
كيف يمكن نقض هذه الحجة؟
دون افتراض نظرية السلف المشترك فإن النتيجة الأرجح هي أن تسلسل بروتين سيتوكروم سي لمختلف الكائنات سيكون مختلفاً كثيراً بين الأنواع (و هذا غير صحيح) و في تلك الحالة فإن تحليل النشوء والتطور سيكون مستحيلاً و سيكون ذلك دليلاً قوياً لعدم ارتباط الأنساب وأصل الأنواع و سيهدم أساس هذه الحجة في حال تم برهنة أن بروتين سيتوكروم سي المحدد لكل نوع يصبح فعالاً على وجه الحصر في الكائن الحي الخاص به فقط أي لا يوجد سيتوكروم فعال من كائن ما فعّال و وظيفي في كائن آخر.
الخلاصة:
الأدلة المقدمة أعلاه تبرهن أن البروتينات الوظيفية الكلية الوجود كالسيتوكروم سي يوجد لها عدد هائل من التسلسلات المعادلة و الموازية و التي يمكنها تشكيل ذلك البروتين في أي كائن حي و لذلك السبب الوحيد لتماثل مثل هذه التسلسلات البروتينية بين كائنان مختلفان في ظل غياب ضرورة وظيفية لهذا التشابه هو الوراثة فعندها يمكننا أن نستنتج بثقة أن الكائنان مرتبطان بالنسب و تماثل تسلسلاتهما ليس اختباراً لنظرية السلف المشترك فحسب بل حسماً لمبدأ الوراثة و أخيراً فإن التماثل ليس مقيداً بالبروتين الكلي الوجود هذا فقد تماثلت كل البروتينات كلية الوجود التي تمت مقارنتها بين الإنسان والشمبانزي بشكل كبير.
مصدر 1 ، مصدر 2 ، مصدر 3 ، مصدر 4 ، مصدر 5</a
اقرأ أيضًا: دراسة جزيئية توضح كيفية تطور الرؤية الملونة عند البشر و أسلافهم
